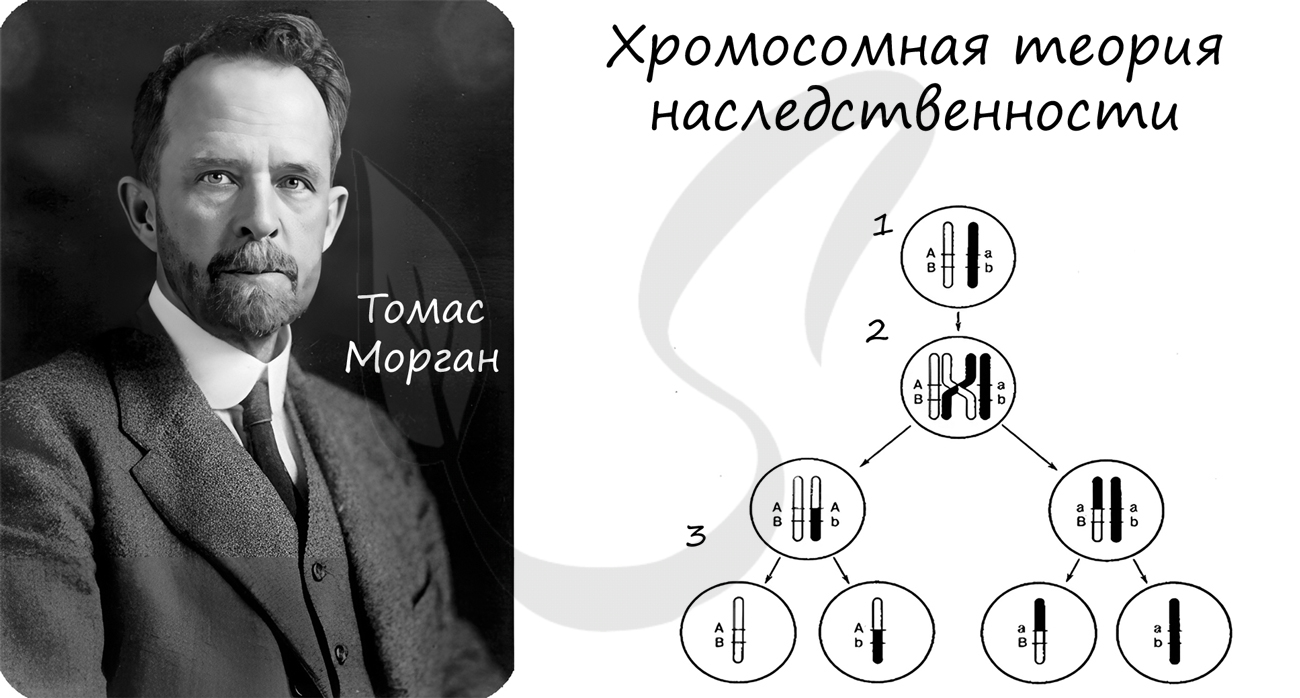
Хромосомная теория наследственности
Концепция данной теории заключается в том, что передача наследственной информации в ряду поколений осуществляется путем передачи хромосом, в которых в определенной линейной последовательности расположены гены.
Данная теория была сформулирована в начале XX века. Значительный вклад в ее развитие внес американский генетик Томас Морган.
Рекомендую осознать и запомнить следующие положения хромосомной теории:
- Гены расположены в хромосомах в линейном порядке
- Каждый ген занимает в хромосоме определенное место — локус
- Гены, расположенные в одной хромосоме, образуют группу сцепления
- Сцепление генов может нарушаться в результате кроссинговера
- Частота кроссинговера между генами прямо пропорциональна расстоянию между ними
- Расстояние между генами измеряется в морганидах (1 морганида — 1% кроссинговера)
Группы сцепления
В предыдущей статье были раскрыты суть и применение в задачах III закона Менделя, закона независимого наследования,
в основе которого лежат гены, расположенные в разных хромосомах. Но что если гены лежат в одной хромосоме? Такие гены образуют группу сцепления, в этом
случае говорят о сцепленном наследовании.
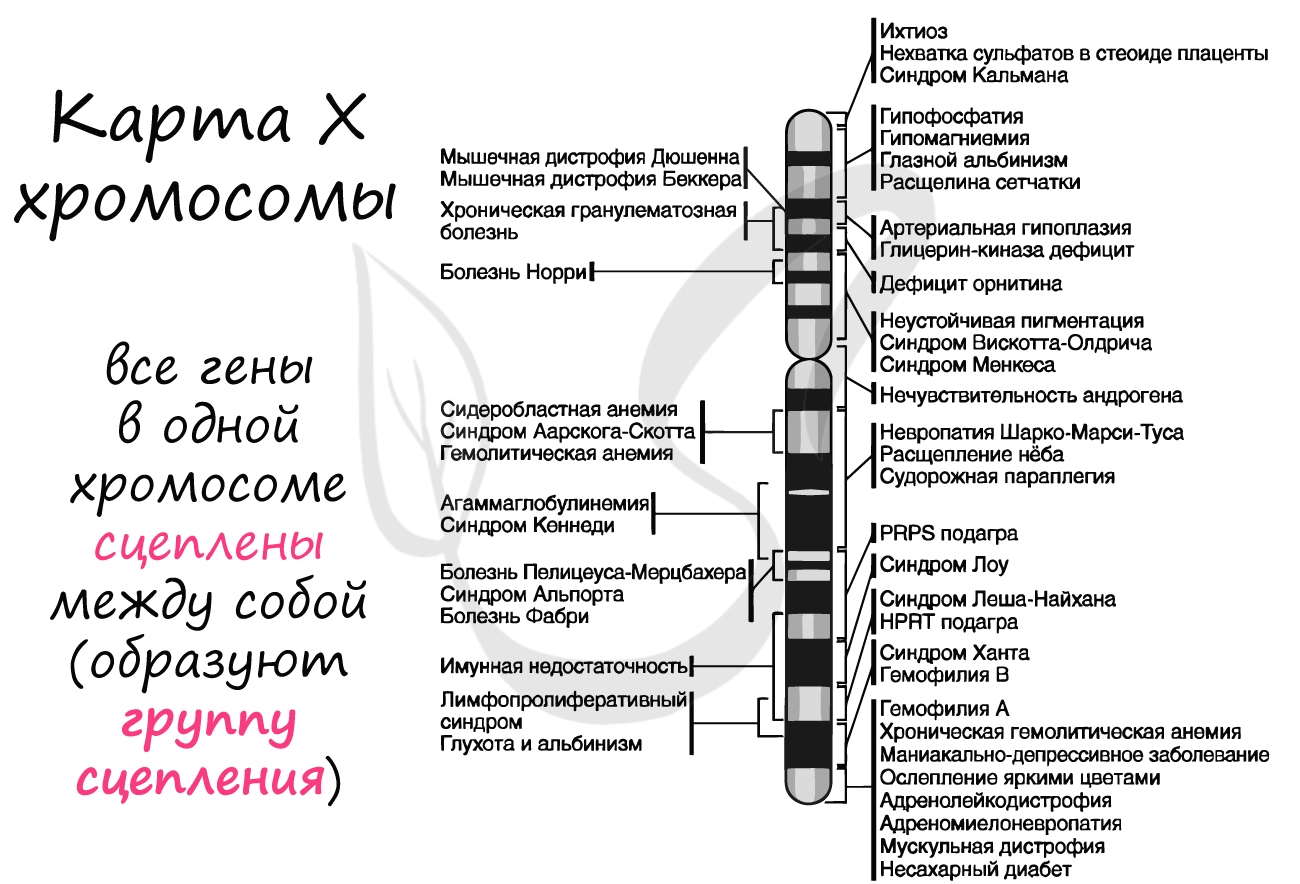
Группа сцепления — совокупность всех генов, расположенных в одной хромосоме, вследствие чего они наследуются совместно. Число групп сцепления равно гаплоидному набору хромосом: у женщины 23 группы сцепления (23 пара —
половые хромосомы XX), а у мужчины — 24 группы сцепления (X и Y представляют собой две отдельные группы).
Сцепление генов
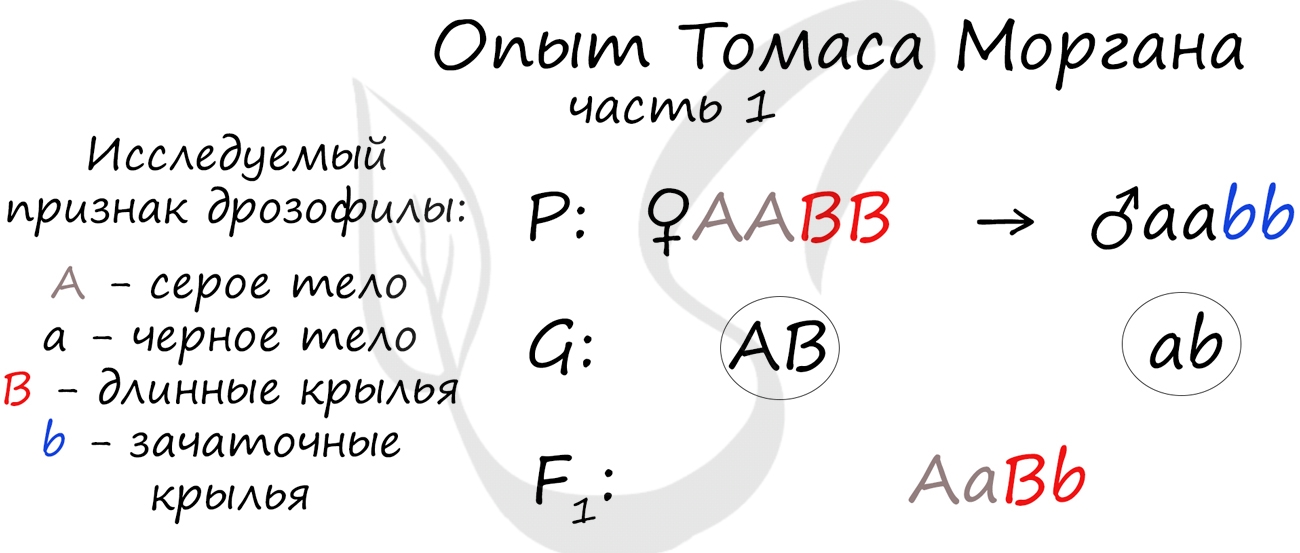
Томас Морган в своих экспериментах изучал наследование признаков плодовых мушек дрозофил: серый (A) — черный (a) цвет тела,
длинные (B) — зачаточные (b) крылья. В первом эксперименте Морган скрестил чистые линии плодовых мушек: серых с длинными
крыльями (AABB) и черных с зачаточными (aabb).
Только что вы видели первый закон Менделя (единообразия) в действии, правда, в несколько ином варианте — при дигибридном
скрещивании. Но суть та же: в первом поколении все особи получаются единообразны по исследуемому признаку, с генотипом
AaBb — с серым телом и длинными крыльями.
Далее Морган применил анализирующее скрещивание. Полученную в первом поколении дигетерозиготу (AaBb) он скрестил с черной особью с
зачаточными крыльями (aabb). Результат весьма удивил Моргана и его коллег: помимо потомства с ожидаемыми фенотипами
(серое тело + длинные крылья, черное тело + зачаточные крылья) были получены особи со смешанными признаками.

Потомство со смешанными признаками подразумевает под собой особи Aabb (серое тело + зачаточные крылья) и aaBb (черные тело +
длинные крылья). Но откуда они могли взяться, если гены A и B находятся в одной хромосоме? Значит, образовались еще какие-то дополнительные гаметы, помимо AB и ab?
Объясняя полученные в потомстве фенотипы, которые содержали смешанные признаки, Томас Морган пришел к выводу, что
между гомологичными хромосомами произошел кроссинговер, в результате которого образовались гаметы Ab, aB — кроссоверные
гаметы.
Очевидно, что в данном случае расстояние между генами A и B было 17 морганид, так как каждой кроссоверной гаметы (соответственно и особей) образовалось
по 8.5%. Не забывайте, что процент кроссинговера равен расстоянию между генами. Поскольку расстояние было 17 морганид = 17%, то на каждую из кроссоверных гамет приходится половина — 8.5%
Пример решения генетической задачи №1
«Катаракта и полидактилия у человека обусловлены доминантными аутосомными генами, расположенными в одной хромосоме.
Гены полностью сцеплены. Какова вероятность родить здорового ребенка в семье, где муж нормален, жена гетерозиготна
по обоим признакам, мать жены также страдала обеими аномалиями, а отец был нормален».
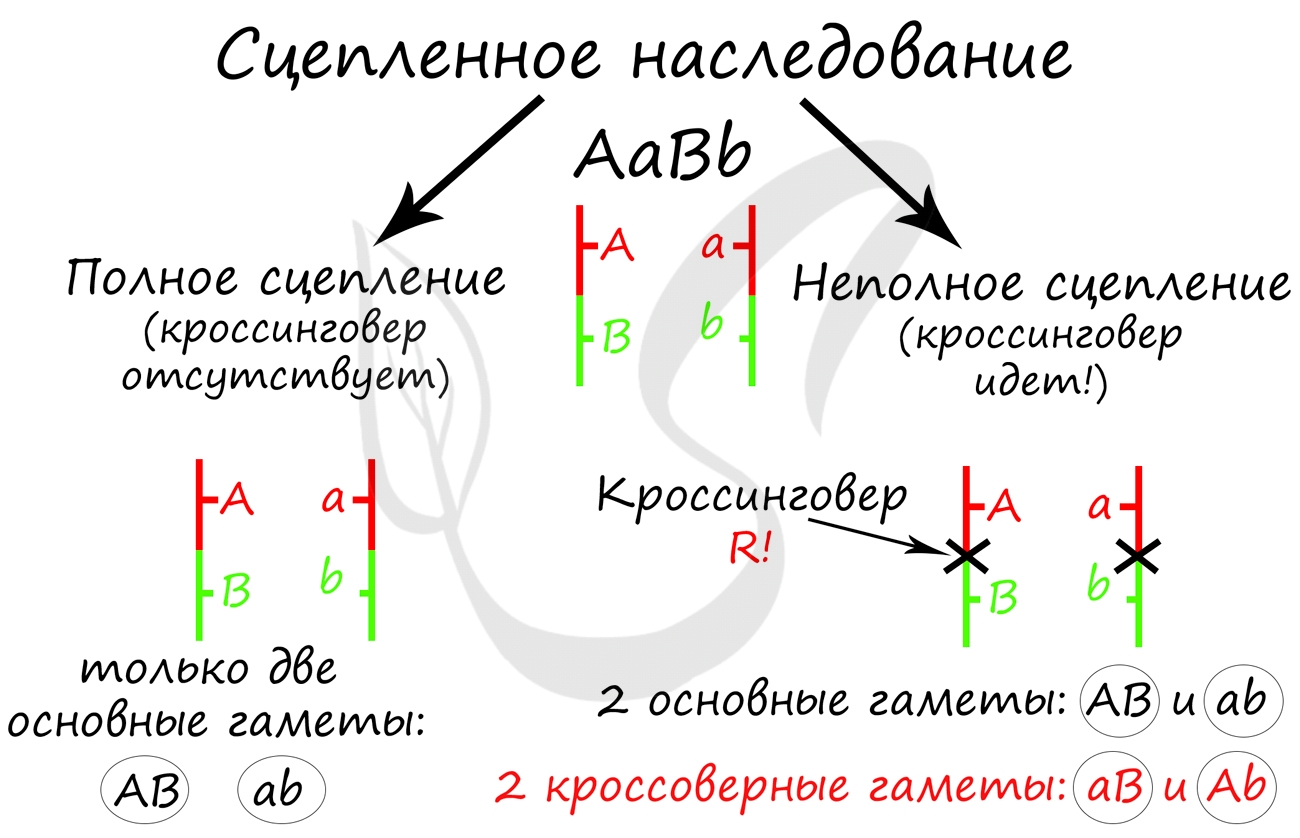
Очень важно обратить внимание на то, что «гены полностью сцеплены» — это говорит об отсутствии кроссинговера, и то, что
мы заметили это, обеспечивает верное решение задачи.

Самое главное, что вам следует усвоить: поскольку гены полностью сцеплены (кроссинговер отсутствует), женщина с генотипом AaBb может образовать только два типа гамет — AB, ab. Кроссоверные гаметы (Ab, aB) не образуются. Всего возможных генотипов потомков получается два, из которых
здоров только один — aabb. Шанс родить здорового ребенка в такой семье ½ (50%).
Пример решения генетической задачи №2
«Гены доминантных признаков катаракты и эллиптоцитоза локализованы в 1-й аутосоме. Гены неполностью сцеплены. Женщина, болеющая
катарактой и эллиптоцитозом, отец которой был здоров, выходит замуж за здорового мужчину. Определите возможные фенотипы потомства и вероятность рождения
больного обеими аномалиями ребенка в этой семье».
Ключевые слова в тексте этой задачи, на которые следует обратить внимание: «гены неполностью сцеплены». Это означает, что между ними
происходит кроссинговер.
Генотип женщины остается неясен из текста задачи. Раз она больна, то он может быть: AaBb, AABB, AABb, AaBB. Однако в тексте дано то, что развеет
сомнения: «отец которой был здоров». Если ее отец был здоров, то его генотип был aabb, значит он передал дочери гамету ab. Теперь
становится очевидно, что генотип дочери AaBb — она дигетерозиготна.

В данном случае между генами A и B произошел кроссинговер, их сцепление нарушилось. В результате образовались кроссоверные гаметы
Ab, aB — которые привели к образованию особей с со смешанными признаками (Aabb, aaBb). Вероятность рождения в этой семье ребенка,
больного обеими аномалиями, составляет ¼ (25%).
Наследование, сцепленное с полом
Половые хромосомы X и Y определяют пол человека. Генотип XX характерен для женщин, а XY — для мужчин. Мужская Y-хромосома
не содержит аллелей многих генов, которые есть в X-хромосоме, вследствие этого наследственными заболеваниями, сцепленными с
полом, чаще болеют мужчины.
Природа, несомненно, бережет женских особей. Женщины имеют две гомологичные хромосомы XX, и если ген наследственного заболевания
попал в одну из X-хромосом, то чаще всего в другой X-хромосоме окажется «здоровый» ген, доминантный, которой подавит действие
рецессивного гена. С генетической точки зрения, женщина будет носительницей заболевания, может его передать по поколению, но
сама болеть не будет.
У мужчин если ген заболевания оказался в X-хромосоме, то не проявиться он не может. Именно по этой причине мужчины чаще
страдают дальтонизмом, гемофилией и т.д.
Не у всех организмов особь мужского пола характеризуется набором хромосом XY, а женского — XX. У пресмыкающихся, птиц,
бабочек женские особи имеют гетерогаметный пол- XY, а мужские — XX. То же самое относится к домашним курам: петух — XX, курица — XY.
Решим несколько задач по теме наследования, сцепленного с полом. Речь в них будет идти о сцепленных с полом признаками —
признаками, гены которых лежат не в аутосомах, а в гетеросомах (половых хромосомах).
Пример решения генетической задачи №3
«Рецессивный ген дальтонизма располагается в X-хромосоме. Женщина с нормальным зрением (отец был дальтоник) выходит замуж
за мужчину с нормальным зрением, отец которого был дальтоником. Определите возможные фенотипы потомства».
Подробности о родословной важны и помогают заполнить белые пятна. Если отец женщины был дальтоником (XdY), то
очевидно, что он передал ей хромосому Xd, так как от отца дочери всегда передается X-хромосома. Значит женщина
гетерозиготна по данному признаку, а у мужчины возможен лишь один вариант здорового генотипа — XDY. То, что его
отец был дальтоником несущественно, ведь отец всегда передает сыну Y-хромосому.

Возможные фенотипы потомства:
- XDXD, XDXd — фенотипически здоровые девочки
- XDY — здоровый мальчик
- XdY — мальчик, который болен дальтонизмом
Пример решения генетической задачи №4
«Гипоплазия зубной эмали наследуется как сцепленный с X-хромосомой доминантный признак, шестипалость — как аутосомно-доминантный.
В семье, где мать шестипалая, а у отца гипоплазия, родился пятипалый здоровый мальчик. Напишите генотипы всех членов семьи по данным
признакам. Возможно ли у них рождение ребенка с двумя аномалиями одновременно?»
Ответ на вопрос: «Каковы генотипы матери и отца?» — лежат в потомстве. Пятипалый здоровый мальчик имеет генотип aaXbY.
Чтобы сформировался такой генотип, от матери должна прийти гамета aXb, а от отца — aY. Выходит, что единственно возможный генотип
матери — AaXbXb, а генотип отца — aaXBY.

Рождение ребенка с двумя аномалиями возможно — AaXBXb, вероятность такого события ¼ (25%).
Пример решения генетической задачи №5
«Рецессивные гены, кодирующие признаки дальтонизма и гемофилии, сцеплены с X-хромосомой. Мужчина с нормальным цветовым зрением и гемофилией женится на здоровой женщине, отец которой был дальтоником, но не гемофиликом. Известно, что мать женщины была гомозиготна по исследуемым признакам. Какое потомство
получится от брака их дочери со здоровым мужчиной?»
Генотип мужчины вопросов не вызывает, так как единственный возможный вариант — XhDY. Генотип женщины
дает возможность узнать ее отец (XHdY), который передал ей гамету XHd (отец всегда передает
дочке X хромосому, а сыну — Y), следовательно, ее генотип — XHDXHd
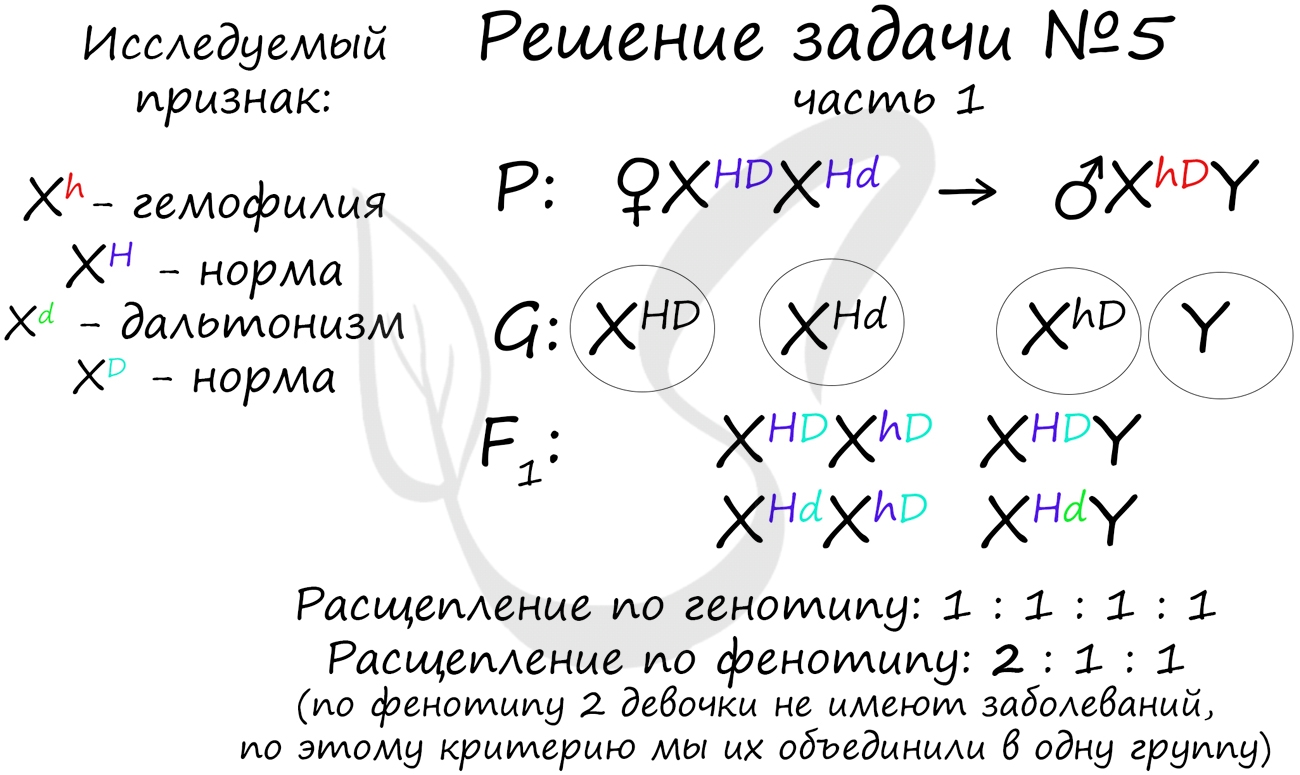
Как оказалось, возможны два варианта генотипа дочери: XHDXhD, XHdXhD.
Генотип здорового мужчины XHDY. Следуя логике задачи, мы рассмотрим два возможных варианта брака.
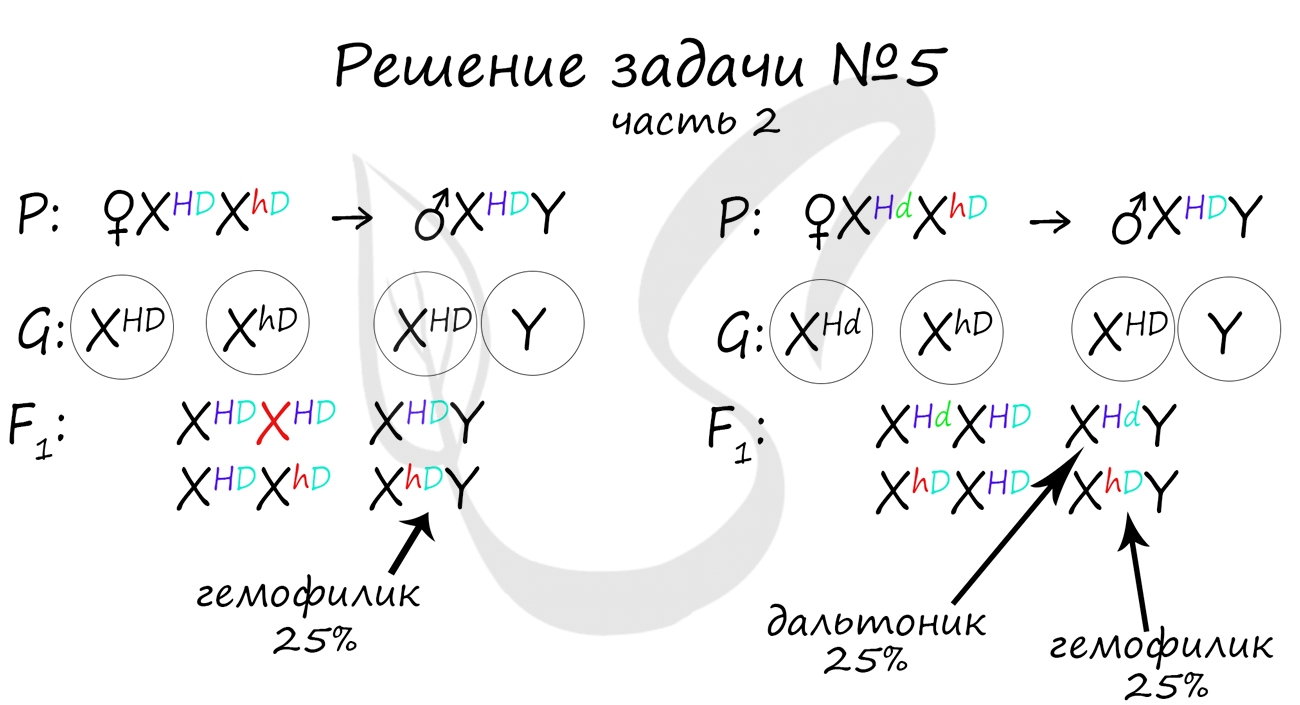
Не забывайте, что на экзамене схема задачи не является ответом. Ответ начинается только после
того, как вы напишите слово «Ответ: …». В ответе должны быть указаны все фенотипы потомства, их описание, что возможно
покажется рутинными при большом числе потомков, но весьма приятным, если вы верно решили задачу и получили за нее заслуженные
баллы 
© Беллевич Юрий Сергеевич 2018-2023
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение
(в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов
без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования,
обратитесь, пожалуйста, к Беллевичу Юрию.
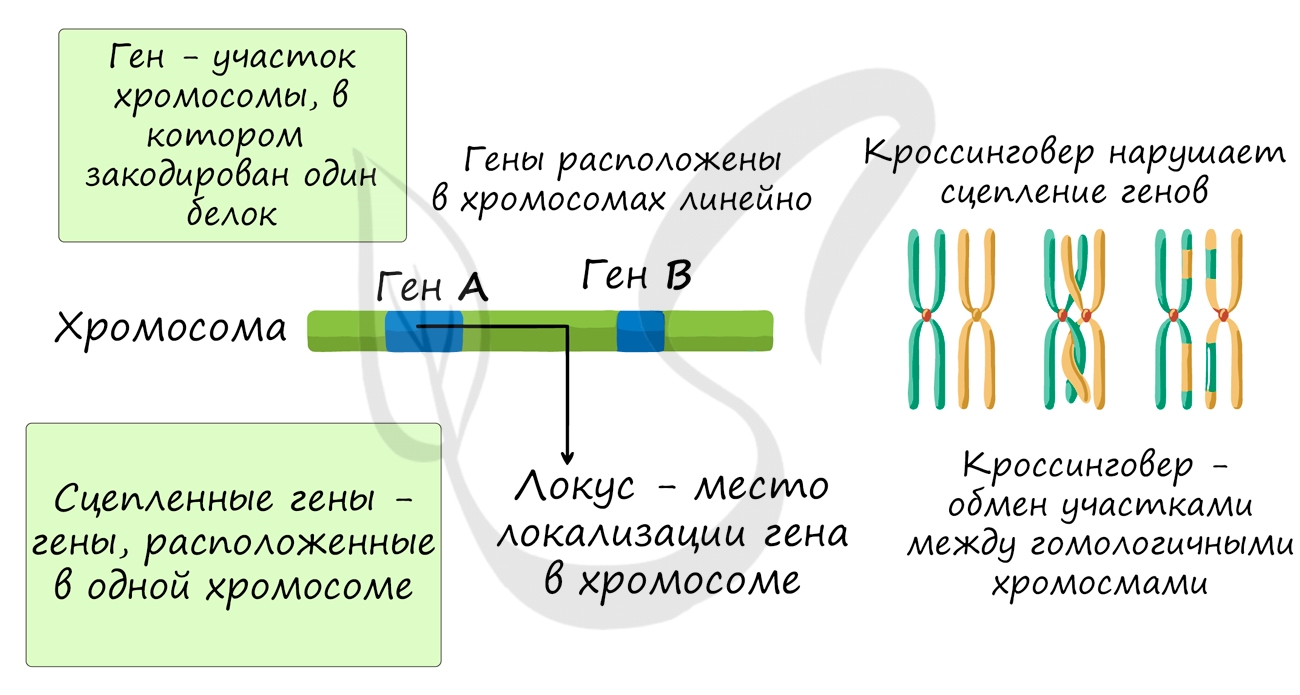
Хромосомная теория наследственности появилась в XX веке. Оказалось, что количество генов в клетке намного больше, чем количество хромосом. Поэтому каждая хромосома содержит сотни или тысячи генов (половая Y-хромосома является исключением.) Каждый ген находится в хромосоме на определённом месте. Это место называется локусом.
Связанные гены (расположенные близко друг к другу) являются сцепленными и наследуются совместно. Всё потому, что хромосомы, а не гены расходятся независимо в гаметы во время мейоза.
Центральная роль хромосом в наследственности была впервые предложена в 1900 г немецким генетиком Карлом Корренсом, в одной из статей, объявляющей о повторном открытии идей Менделя. Вскоре после этого появились наблюдения, что гомологичные хромосомы во время мейоза расходятся в разные гаметы. Хромосомная теория наследственности была впервые независимо друг от друга сформулирована Уолтером Саттоном в 1903 г и Теодором Бовери в 1902 г.

Хромосомная теория наследственности: Морган впервые соотнёс наследование признаков с половыми хромосомами
Подробное экспериментальное доказательство и объяснение хромосомная теория наследственности получила от Томаса Ханта Морган и его сотрудников. Они доказали, что гены расположены в хромосомах линейно и наследуются сцепленно и что это сцепление может нарушать кроссинговер.
В 1910 г, изучая плодовую мушку дрозофилу обыкновенную, или дрозофилу фруктовую (Drosophila melanogaster), Морган обнаружил мутировавшего самца мухи с белыми глазами вместо красных. Он немедленно приступил к её изучению, желая узнать, будет ли эта черта наследоваться менделеевским способом.

Сначала он скрестил мутантного самца с нормальной красноглазой самкой, чтобы посмотреть, какой из двух признаков доминирует. У всего потомства F1 были красные глаза, поэтому Морган заключил, что красный цвет глаз доминирует над белым.
Скрещивание особей поколения F1
Следуя экспериментальной процедуре, установленной Менделем, Морган скрестил особей из поколения F1 между собой. Из 4252 особей поколения F2 782 (18%) имели белые глаза. Хотя соотношение красно- и белоглазых дрозофил было больше чем 3:1, оно послужило доказательством того, что цвет глаз наследуется независимо. Однако нечто в исходе скрещивания было странным и непредсказуемым – все белоглазые мухи поколения F2 были самцами.

Морган стал искать объяснение такому результату. Одна из его версий была следующей – белоглазых самок просто не существует, такие особи по какой-то причине оказываются нежизнеспособными. Чтобы проверить эту идею он скрестил красноглазых самок поколения F1 с белоглазыми самцами.
И получил соотношение полов с белыми и красными глазами в соотношении 1:1:1:1, как и предсказывал закон независимого наследования. Оказалось, что белоглазые самки существуют. Тогда Морган обратился к природе половых хромосом. Это и было первым опытным доказательством хромосомного наследования .
В 1910 г появилась публикация об этой мутации, в ней Морган отметил, что характер её наследования совпадает с наследованием половых хромосом дрозофилы. Вскоре было описано ещё две сцепленных с полом мутации, и при изучении их совместного наследования Морган пришёл к заключению, что гены должны быть организованы на хромосоме линейно, и их сцепленное наследование может нарушаться из-за кроссинговера.
Хромосомная теория наследственности: основные положения:
- Гены находятся в хромосомах.
- Хромосомы содержат неодинаковое число разных генов, набор генов негомологичных хромосом уникален.
- Гены в хромосоме расположены в линейной последовательности.
- Аллели одного гена занимают одинаковые локусы в гомологичных хромосомах.
- Гены одной хромосомы образуют группу сцепления, то есть наследуются преимущественно сцепленно (совместно), благодаря чему происходит сцепленное наследование некоторых признаков. Число групп сцепления равно гаплоидному числу хромосом данного вида (у гомогаметного пола) или больше на 1 (у гетерогаметного пола). У человека — 23 группы сцепления у женщин и 24 у мужчин.
- Сцепление нарушается в результате кроссинговера, частота которого прямо пропорциональна расстоянию между генами в хромосоме (поэтому сила сцепления находится в обратной зависимости от расстояния между генами).
- Каждый биологический вид характеризуется определённым набором хромосом — кариотипом.
- За единицу расстояния между сцепленными генами принята 1 морганида – расстояние, на котором кроссинговер происходит с вероятностью 1%.
Одна из первых открытых сцепленных с неполовыми хромосомами пар генов
Хромосомная теория наследственности объясняет передачу генов не только сцепленных с половыми хромосомами. У душистого горошка ген окраски цветка (фиолетовый против красного) и ген формы пыльцевого зерна (круглые или удлинённые) переносятся в одной хромосоме. Поэтому аллели этих генов наследуются вместе.
Гетерозиготные растения душистого горошка имеют фиолетовые цветки и удлинённые пылинки. Аллели, отвечающие за фиолетовую окраску и удлинённую форму пыльцы лежат в одной гомологичной хромосоме, а отвечающие за красный цвет и круглую форму – в другой. Значит, две гаметы этого растения будут содержать либо аллели с фиолетовым цветом и овальной формой, либо с красной окраской и круглой формой пыльцевого зерна.
Такой тип наследования не соответствует независимому наследованию, поскольку окраска цветка и форма не отделяются во время мейоза.
Как связь двух генов влияет на наследование?
Чтобы увидеть, как связь между генами влияет на наследование двух разных признаков, давайте рассмотрим еще один из экспериментов Моргана с дрозофилами. В этом случае будем следить за наследованием окраски тела и размеров крыльев мух.
Дикие плодовые мушки имеют серые тела и крылья нормального размера. Вдобавок к этим мухам Морган успел обзавестись мутантными особями с черными телами и крыльями намного меньше обычных – рудиментарными. Мутантные аллели являются рецессивными по отношению к аллелям дикого типа. Во время изучения наследования этих двух генов, Морган провел скрещивание, показанное на рисунке ниже.
Сначала он скрестил чистые линии этих мух с серым телом и нормальными крыльями (ААВВ) и с чёрным телом и зачаточными крыльями (аавв). Все гибриды первого поколения в соответствии с законом единообразия были серыми с нормальными крыльями (АаВв).

Скрестив между собой гибридов первого поколения (АаВв х АаВв) при полном сцеплении генов АВ и ав, мы бы получили соотношение по фенотипу 3:1.
Морган же провёл анализирующее скрещивание. Он скрестил дигетерозиготную (АаВв) самку из первого поколения с рецессивным дигомозиготным самцом (аавв). В потомстве было получено по 41,5 % особей с серым телом, нормальными крыльями и черным телом, зачаточными крыльями, а также по 8,5 % мух с серым телом, зачаточными крыльями и черным телом, нормальными крыльями.

Если бы гены, определяющие цвет тела и развитие крыльев, находились в разных парах хромосом, соотношение фенотипических классов было бы равным — по 25 %. Но этого не наблюдалось, значит, гены находятся в гомологичных хромосомах и наследуются сцепленно.
Несмотря на сцепление генов самка произвела не два, а четыре типа гамет, но гамет с исходным сочетанием (АВ, ав) было намного больше (83%), чем новых – Ав, аВ (17%).
Таким образом, Морган пришел к выводу, что цвет тела и размер крыла обычно наследуются вместе в специфических (родительских) комбинациях, потому что гены для активации этих признаков находятся рядом друг с другом на одной хромосоме.
Однако обе комбинации черт, не замеченные в родительском поколении, тоже были получены в экспериментах Моргана. Он предположил, что аллели цвета тела и размера крыла не всегда связаны — сцепление неполное. Чтобы проверить эту гипотезу, нужно было дальше исследовать генетическую рекомбинацию, применяя выведение потомства с комбинациями, отличающихся признаков, что были найдены у родителей.
Хромосомная теория наследственности и генетическая рекомбинация
Мейоз и случайное оплодотворение порождают генетические вариации среди потомства у организмов, размножающихся половым путем. О независимом наследовании Мендель узнал из скрещиваний, в которых он следил за двумя признаками гороха. Он увидел, что некоторые потомки имеют черты, которые не совпадают ни с одной из родительских. Скрещивая растения с жёлтыми круглыми семенами с растениями с зелёными морщинистыми, он получил также жёлтые морщинистые и зелёные круглые семена (рекомбинантные, или кроссоверные).
Но половина потомства унаследовала фенотип, который соответствует одному из родительских. Когда 50% всего потомства являются рекомбинантами, как в данном примере говорят, что существует частота рекомбинации равная 50%. Частота рекомбинации в 50% также наблюдаются для любых двух генов, расположенных на разных хромосомах.
Теперь давайте вернемся в «летную комнату» Моргана, чтобы посмотреть, как можно проиллюстрировать результаты тесткросса. Напомним, что большинство отпрысков по окраске тела и размерам крыла имел родительские фенотипы.
Это дало возможность предположить, что два гена были в одной хромосоме. Появления родительских типов в количестве больше 50% указывает на то, что гены связаны. Около 17% потомства, однако, были рекомбинантами, значит имел место кроссинговер.
При полном сцеплении в результате анализирующего скрещивания получается только 2 фенотипа в соотношении 1:1.

Столкнувшись с этими результатами, Морган предложил, что какой-то процесс должен иногда прерывать физическое соединение между определенными аллелями генов одной хромосомы. Следующие эксперименты показали, что этот процесс, в настоящее время называемый кроссинговером, влияет на рекомбинацию связанных генов.
Гомологичные хромосомы спариваются во время профазы мейоз I (конъюгация), набор ферментов организует обмен (кроссинговер) сегментов отцовской и материнской хромосом. По сути, конечные части двух хроматид меняются местами каждый раз, когда происходит их пересечение.

Вариации для естественного отбора
Физическое поведение хромосом во время мейоза способствует генерации вариаций в потомстве. Каждая пара гомологичных хромосом выстраивается независимо от других пар во время метафаза I, во время профазы I смешиваются и сочетаются части материнского и отцовского гомологов. Это одна из причин наследственной изменчивости организмов – комбинативная.
Причины комбинативной изменчивости:
- перетасовка аллелей генов во время кроссинговера;
- независимое расхождение хромосом в процессе мейоза;
- случайность встречи гамет во время оплодотворения и значит случайный набор хромосом.
Отображение расстояния между генами с помощью данных по рекомбинации
Хромосомная теория наследственности продолжала развиваться. Открытие связанных генов и их рекомбинации привело одного из учеников Моргана Альфреда Х. Стертеванта к открытию метода построения генетической карты – упорядоченного «списка» генетических локусов внутри определенной хромосомы.
Стертевант предположил, что процент рекомбинации и частота рекомбинации зависят от расстояния между генами на хромосоме. Он подумал, что пересечение является случайным событием, с шансом перехода примерно равным во всех точках вдоль хромосомы. Основываясь на этих предположениях, Стертевант сделал вывод, что чем дальше друг от друга находятся два гена, тем выше вероятность того, что кроссовер будет происходить между ними, и поэтому будет выше частота рекомбинации.
Его рассуждения были просты: чем больше расстояние между двумя генами, тем больше точек, в которых может произойти пересечение. С помощью данных по рекомбинации от различных скрещиваний плодовой мухи, Стертевант присваивал относительные позиции генам на хромосоме.
Генетическая карта, основанная на частотах рекомбинации, называется карта связей.

Расин выразил расстояния между генами в единицах карты, определяющие одну карту блок как эквивалент к частоте рекомбинации 1%. В честь Т. Моргана единица измерения расстояния между генами получила название морганиды или, что то же самое, сантиморганиды (сМ).
Морганида (сантиморганида, сМ) — это генетическое расстояние, на котором кроссинговер происходит с вероятностью 1 %.
На практике интерпретация данных рекомбинации сложнее, чем в этом примере. Некоторые гены на хромосоме настолько далеки друг от друга, что сцепления между ними практически никогда не происходит. Наблюдаемая частота повторных комбинаций в скрещиваниях с участием двух таких генов может иметь максимальное значение 50% — результат неотличим от того, когда гены расположены на разных хромосомах. В этом случае физическая связь между генами на одной хромосоме не отражается в результатах генетических скрещиваний.
Несмотря на то, что они находится одной хромосоме, аллели таких генов разбираются по отдельности, как будто они на разных хромосомах. По крайней мере, два из генов гороха, которые изучал Мендель, находятся на одной и той же хромосоме, но расстояние между ними настолько велико, что связь при генетическом скрещивании не наблюдается. Следовательно, оба гена в экспериментах Менделя вели себя так, как будто они находились на разных хромосомах.
Как высчитать частоту кроссинговера?
Кроссинговер между конкретными сцепленными генами происходит с определенной вероятностью (частотой). Для расчета частоты кроссинговера (rf, от англ. recombination frequency — частота рекомбинации) можно пользоваться следующей формулой:
rf =сумма кроссоверных гамет (особей) /общее количество гамет (особей) х100 %.
Таким образом, между генами А и В, контролирующими цвет тела и длину крыльев дрозофилы, кроссинговер происходит с частотой: rfАВ = 17 %.
Второй способ: если известно расстояние между генами на хромосоме. Рассмотрим частоту кроссинговера между генами на рисунке:

А-В — 5%; А-С — 12%; В-С — 7%.
Запись и решение задач на сцепленное наследование и кроссинговер
Хромосомная теория наследственности — одна из сложнейших тем генетики. Задачи и примеры скрещивания на сцепленное наследование удобнее и нагляднее записывать не генным, а хромосомным способом.
Автор материалов этого раздела: Дашков М.Л. Источник: интернет-сайт dashkov.by
1. Серый цвет тела у дрозофилы доминирует над жёлтым, красные глаза — над гранатовыми. Гены, отвечающие за эти признаки, локализованы в первой паре хромосом и находятся на расстоянии 44 морганиды. Скрестили чистые линии серотелых мух с гранатовыми глазами и жёлтотелых с красными глазами. Из полученных гибридов выбрали самку и подвергли её анализирующему скрещиванию. Каким будет процентное соотношение фенотипических классов в потомстве?
- Введём обозначения генов, укажем расстояние между ними:
А – серое тело;
а – жёлтое тело;
B – красные глаза;
b – гранатовые глаза;
rfAB = 44% (44 морганиды).
- Установим генотипы родительских форм. Особи чистой линии серотелых мух с гранатовыми глазами имеют генотип Ab//Ab, чистой линии жёлтотелых мух с красными глазами – генотип aB//aB.
- В результате скрещивания чистых линий было получено первое гибридное поколение:


- Следовательно, самка, которую подвергли анализирующему скрещиванию, имеет генотип Ab//aB.
Запишем анализирующее скрещивание, учитывая, что у дигетерозиготной самки образуется два типа некроссоверных гамет (в равном соотношении) и два типа кроссоверных гамет (также в равном соотношении). Для удобства выделим кроссоверные гаметы и особи звёздочками (*):


- Рассчитаем процентное соотношение особей в Fа.
Расстояние между генами – 44 морганиды, значит, кроссинговер между ними протекает с вероятностью 44%. Следовательно, общее количество кроссоверных особей составит 44% (а особей каждого фенотипического класса – по 22%). Общее количество некроссоверных особей равно: 100% – 44% = 56% (т.е. каждый фенотипический класс – по 28%).
Ответ: в потомстве будет получено по 28% особей с серым телом, гранатовыми глазами и с жёлтым телом, красными глазами, а также по 22% особей с серым телом, красными глазами и с жёлтым телом, гранатовыми глазами.
- У одного из видов растений рассечённые листья доминируют над цельными, а синяя окраска цветков — над розовой. В результате анализирующего скрещивания было получено потомство четырёх фенотипических классов:
1) 133 растения с рассечёнными листьями и синими цветками;
2) 362 растения с рассечёнными листьями и розовыми цветками;
3) 127 растений с цельными листьями и розовыми цветками;
4) 378 растений с цельными листьями и синими цветками.
Затем растения первого фенотипического класса скрестили с растениями второго фенотипического класса. Сколько процентов полученных гибридов будут иметь рассечённые листья и розовые цветки? Цельные листья и синие цветки?
- Введём обозначения генов:
А – рассечённые листья;
а – цельные листья;
B – синие цветки;
b – розовые цветки.
- В результате анализирующего скрещивания было получено четыре фенотипических класса в неравном соотношении. Из этого можно сделать следующие выводы:
1) Гены, определяющие тип листьев и окраску цветков, наследуются сцепленно.
2) Анализируемая особь продуцировала 4 типа гамет, т.е. была дигетерозиготной.
3) Потомки первого и третьего фенотипических классов являются кроссоверными (их примерно поровну и намного меньше, чем особей второго и четвёртого фенотипических классов). Потомки второго и четвёртого фенотипических классов – некроссоверные (их примерно поровну и намного больше, чем особей первого и третьего фенотипических классов).
4) Частота кроссинговера между генами А и В составляет: rfAB = сумма кроссоверных особей/общее количество особей × 100% = (133 + 127) : (133 + 362 + 127 + 378) × 100% = 260 : 1000 × 100% = 26%.
- Установим генотипы особей первого и второго фенотипических классов. Те и другие унаследовали сцепленные гены ab от рецессивного гомозиготного родителя ab//ab.
Особи первого фенотипического класса имеют рассечённые листья (А) и синие цветки (В), значит, от другого родителя (анализируемого) они унаследовали сцепленные гены AB. Таким образом, особи первого фенотипического класса имеют генотип AB//ab.
Особи второго фенотипического класса имеют рассечённые листья (А) и розовые цветки (b), следовательно, от другого родителя (анализируемого) они унаследовали сцепленные гены Ab. Таким образом, особи второго фенотипического класса имеют генотип Ab//ab.
Запишем скрещивание особей первого и второго фенотипических классов, для удобства выделим кроссоверные гаметы и особи звёздочками (*):


- Для того, чтобы найти долю особей в каждой ячейке решётки Пеннета, нужно умножить друг на друга доли соответствующих гамет.
Следовательно, процентная доля растений с рассечёнными листьями и розовыми цветками составляет: (0,37 × 0,5 + 0,13 × 0,5 + 0,13 × 0,5) × 100% = (0,185 + 0,065 + 0,065) × 100% = 0,315 × 100% = 31,5%.
Процентная доля растений с цельными листьями и синими цветками равна: 0,13 × 0,5 × 100% = 0,065 × 100% = 6,5%.
Ответ: 31,5% гибридов будут иметь рассечённые листья и розовые цветки; 6,5% гибридных растений будут иметь цельные листья и синие цветки.
3.

Сцепленное наследование генов
Необходимо запомнить
ВАЖНО!
Сцепление генов – это совместное наследование генов, расположенных в одной и той же хромосоме. Количество групп сцепления соответствует гаплоидному числу хромосом, то есть у дрозофилы 4. Природу сцепленного наследования объяснил Морган с сотрудниками. В качестве объекта исследования они избрали плодовую муху дрозофилу, которая оказалась очень удобной моделью для изучения данного феномена, так в клетках её тела находится только 4 пары хромосом и имеет место высокая скорость размножения (в течение года можно исследовать более 20-ти поколений). Итак, сцепленными признаками называются признаки, которые контролируются генами, расположенными в одной хромосоме. Естественно, что они передаются вместе в случаях полного сцепления (закон Моргана). Полное сцепление встречается редко, обычно – неполное, из-за влияния кроссинговера (перекрещивания и обмена участками гомологичных хромосом в процессе мейоза). То есть, гены одной хромосомы переходят в другую, гомологичную ей.
Частота кроссинговера зависит от расстояния между генами. Чем ближе друг к другу расположены гены в хромосоме, тем сильнее между ними сцепление и тем реже происходит их расхождение при кроссинговере, и, наоборот, чем дальше друг от друга отстоят гены, тем слабее сцепление между ними и тем чаще возможно его нарушение.

Рис. 1
На рисунке 1 слева: расстояние между генами А и В маленькое, вероятность разрыва хроматиды именно между А и В невелика, поэтому сцепление полное, хромосомы в гаметах идентичны родительским (два типа), других вариантов не появляется.
На рисунке 1 справа: расстояние между генами А и В большое, повышается вероятность разрыва хроматиды и последующего воссоединения крест-накрест именно между А и В, поэтому сцепление неполное, хромосомы в гаметах образуются четырёх типов – 2 идентичные родительским (некроссоверные) + 2 кроссоверных варианта.
Количество разных типов гамет будет зависеть от частоты кроссинговера или расстояния между анализируемыми генами. Расстояние между генами исчисляется в морганидах – единицах расстояния между генами, находящимися в одной хромосоме. 1 морганида соответствует 1 % кроссинговера. Такая зависимость между расстояниями и частотой кроссинговера прослеживается только до 50 морганид. Частота кроссинговера между определенной парой генов – довольно постоянная величина (хотя радиация, химические вещества, гормоны, лекарства влияют на нее; например, высокая температура стимулирует кроссинговер).
Пример, основанный на опытах Моргана

Рис. 2
Гены, расположенные в одной хромосоме, наследуются совместно. Фенотипы А – серое тело, нормальные крылья (повторяет материнскую форму). Б – тёмное тело, короткие крылья (повторяет отцовскую форму). В – серое тело, короткие крылья (отличается от родителей). Г – тёмное тело, нормальные крылья (отличается от родителей). В и Г получены в результате кроссинговера в мейозе.
Если скрестить мушку дрозофилу, имеющую серое тело и нормальные крылья (на рисунке самка), с мушкой, обладающей тёмной окраской и зачаточными (короткими) крыльями (на рисунке самец), то в первом поколении гибридов все мухи будут серыми с нормальными крыльями (А). Это гетерозиготы по двум парам аллельных генов, причём ген, определяющий серую окраску брюшка, доминирует над тёмной окраской, а ген, обусловливающий развитие нормальных крыльев, — доминирует над геном недоразвитых крыльев.
При анализирующем скрещивании гибрида F1 с гомозиготной рецессивной дрозофилой (Б) подавляющее большинство потомков F2 будет сходно с родительскими формами.
Это происходит потому, что гены, отвечающие за серое тело и нормальные крылья– сцепленные гены, также как и гены, отвечающие за тёмное тело и короткие крылья, т. е. они находятся в одной хромосоме. наследование сцепленных генов называют сцепленным наследованием.
Сцепление может нарушаться. Это доказывает наличие особей В и Г на рисунке, т. е. если бы сцепление не нарушалось, то этих особей бы не существовало, однако они есть. Это происходит в результате кроссинговера, который и нарушает сцепленность этих генов.

Рис. 3
На рисунке 3 опыт Моргана отображен подробно. Несцепленное наследование: два гена находятся в разных хромосомах, гетерозигота с равной вероятностью дает четыре типа гамет:

Рис. 4
Сцепленное наследование: два гена находятся в одной хромосоме.
а) При полном сцеплении гетерозигота дает только два типа гамет.
б) При неполном сцеплении гетрозигота дает четыре типа гамет, но не с равной вероятностью.

Рис. 5
На вышесказанном строится хромосомная теория наследственности Моргана:
1. Гены находятся в хромосомах и расположены в линейной последовательности на определенных расстояниях друг от друга.
2. Гены, расположенные в одной хромосоме, составляют группу сцепления. Число групп сцепления равно гаплоидному числу хромосом. Признаки, гены которых находятся в одной хромосоме, наследуются сцепленно (т. е. в тех же сочетаниях, в которых они были в хромосомах исходных родительских форм).
3. Новые сочетания генов, расположенных в одной паре хромосом, могут возникать в результате кроссинговера в процессе мейоза. Частота кроссинговера зависит от расстояния между генами.
4. Учитывая линейное расположение генов в хромосоме и частоту кроссинговера как показателя расстояния между генами, можно построить карты хромосом. За единицу расстояния между генами принята частота кроссинговера равная 1 % (морганида, сантиморган, сМ).
Задачи на нахождение расстояния между генами
Решение задачи на определение вероятности рождения здоровый и больных детей
Тема
6. Сцепленное наследование
признаков и
кроссинговер
Геном
любого организма содержит тысячи и
десятки тысяч генов, в то время как его
кариотип представлен лишь небольшим
количеством хромосом — материальных
носителей генов. В связи с этим одна и
та же хромосома должна содержать не
один, а множество генов. Совокупность
таких генов, лежащих в одной хромосоме,
называют группой сцепления, а сами гены
называются сцепленными. Сцепленные
гены в ряду поколений передаются вместе,
единым блоком. Соответственно, признаки,
гены которых находятся в одной хромосоме,
так же будут сцепленными, и также будут
передаваться вместе. Число групп
сцепления равно гаплоидному набору
хромосом. Исключением являются высшие
организмы, у которых есть гетерогаметный
пол с XY-хромосомами,
поскольку последние представляют собой
две разные группы сцепления. Так, у
человека диплоидный набор равен 46
хромосомам, а групп сцепления 24: 22
соответствуют 22 аутосомам и 2 – X—
и Y-хромосомам.
Совместное
наследование генов ограничивает их
свободное комбинирование во время
гаметогенеза. Однако сцепление генов,
как правило, не бывает полным. Благодаря
обмену идентичными участками гомологичных
хромосом, происходящему в профазе I
мейоза и получившему название перекреста,
или кроссинговера, сцепление нарушается,
чем обеспечивается дополнительный
источник комбинативной изменчивости.
Установить
факт сцепления между двумя генами, а
так же его характер (т.е. является ли оно
полным или неполным), можно либо по
результатам анализирующего скрещивания,
либо по результатам расщепления во
втором поколении. Рассмотрим три случая
анализирующего скрещивания: в первом
случае анализируемые гены локализованы
в разных хромосомах и, как следствие,
проявляют независимое наследование,
во втором –
гены полностью сцеплены, и, наконец, в
третьем случае –
гены сцеплены, но сцепление между ними
не полное.
Скрещивание 1: гены
локализованы в разных хромосомах,
наследуются независимо.
P:
А
B
a
b
a b
a b
GP:
1/4 AB,
1/4 Ab,
1/4 aB,
1/4 ab
ab
F1:
|
1/4 |
1/4 |
1/4 |
1/4 |
|
|
ab |
A a b |
A a b |
a a b |
a a b |
Скрещивание 2: гены
сцеплены, сцепление полное.
P:
A
B
a
b
a
b a b
GP:
1/2 AB,
1/2 ab
ab
F:
|
1/2 |
1/2 |
|
|
ab |
A B a b |
a b a b |
Скрещивание 3: гены
сцеплены, сцепление неполное.
P:
A
B
a
b
a
b a b
GP:
1/4 AB,
1/4
Ab,
1/4 aB,
1/4 ab
ab
F:
|
1/4 |
1/4 |
1/4 |
1/4 |
|
|
ab |
A B a b |
A a b |
a a b |
a a b |
Согласно результатам
первого скрещивания, при независимом
наследовании у дигетерозиготной особи
образуется четыре класса гамет (AB,
Ab, aB,
ab) с одинаковой частотой
— каждого класса примерно по 25%. Как
следствие, и потомки от такого скрещивания
так же будут представлены четырьмя
фенотипическими классами в равном
соотношении 1 : 1 : 1 : 1. При полном сцеплении
генов у дигетерозиготной особи образуется
только два класса гамет (AB
и ab) с родительским
сочетанием аллелей, причем в равном
соотношении. Наконец, при неполном
сцеплении генов у дигетерозиготной
особи образуется четыре класса гамет:
два класса родительских (некроссоверных)
гамет, несущих исходные комбинации
аллелей (AB
и ab), и два класса
кроссоверных, или рекомбинантных
гамет, которые несут хромосомы с
новыми комбинациями аллелей (Ab
и aB). При этом соотношение
таких классов гамет и, как следствие,
фенотипических классов потомства, будет
иным, нежели при независимом наследовании:
частота появления особей родительского
типа будет достоверно выше, чем частота
появления особей кроссоверного типа.
Внутри же одного типа (родительского
или кроссоверного) гаметы (особи), которые
в таком случае называют реципрокными
классами, появляются с одинаковой
частотой.
Частота появления
кроссоверных особей определяется
частотой кроссинговера (rf),
происходящего между двумя сцепленными
генами. Этот показатель рассчитывается
по формуле:
сумма
кроссоверных особей
rf
=
100%
сумма
всех особей
Величина
кроссинговера является мерой расстояния
(силы сцепления) между генами: чем ближе
друг к другу расположены гены в хромосоме,
тем реже между ними происходит
кроссинговер, и, наоборот, чем дальше
друг от друга гены располагаются, тем
чаще между ними возможен перекрест. Из
результатов скрещивания 1
следует, что гены наследуются независимо,
если частота кроссинговера между ними
составляет 50%. Однако при этом гены не
всегда должны быть локализованы в разных
хромосомах: они могут принадлежать к
одной группе сцепления, но находится
на расстоянии более 50 сМ (1сМ = 1%
кроссинговера) друг от друга. Так, гены,
контролирующие окраску цветка и семени
у садового гороха, локализованы в одной
хромосоме на расстоянии 200 сМ.
Таким образом,
разная частота появления особей в
анализирующем скрещивании позволяет:
-
утверждать о том,
что гены сцеплены; -
установить силу
сцепления (частоту кроссинговера) между
этими генами; -
установить частоту
образования кроссоверных гамет и гамет
родительского типа (100% — частота
кроссоверных гамет); -
определить генотип
гетерозиготного родителя (по классам
особей родительского типа, т.е. особей,
возникающих с большей частотой). При
этом следует помнить, что генотип
кроссоверных гамет и, как следствие,
кроссоверных особей, зависит от исходного
положения доминантных аллелей в
гомологичных хромосомах дигетерозиготной
особи. В рассмотренном выше примере
доминантные аллели имеют цис-положение
(состояние «притяжение»). В случае же
транс-положения (состояния «отталкивания»)
доминантных аллелей кроссоверными
будут гаметы с генотипом AB
и ab, которые в предыдущем
варианте являются исходным родительским
типом гамет.
РЕШЕНИЕ ТИПОВЫХ
ЗАДАЧ
Наиболее типичными
генетическими задачами по теме «Сцепленное
наследование и кроссинговер» являются
задачи трех вариантов:
1) задачи на
определение гамет у гетерозиготной
особи при цис- и транс-положении
доминантных аллелей;
2) задачи на
определение частоты кроссинговера
между сцепленными генами по результатам
анализирующего скрещивания;
3) задачи на
определение частоты кроссинговера
между сцепленными генами по результатам
второго поколения.
Из перечисленных
вариантов последний является наиболее
сложным. Для того чтобы определить
частоту кроссинговера по результатам
второго поколения необходимо (в случае
дигетерозиготы типа AaBb):
1) найти долю
рецессивных гомозигот aabb;
2) извлечь
корень квадратный из частоты рецессивных
гомозигот, тем самым, определив частоту
образования гамет типа ab.
Если частота гамет ab
составляет меньше 25%, то эти гаметы
являются гаметами кроссоверного типа
и для определения частоты кроссинговера
необходимо удвоить значение частоты
гаметы ab
(т.к. частота кроссинговера –
это сумма двух кроссоверных гамет, а
они возникают с одинаковой вероятностью).
Если же частота гамет ab
больше 25%, то эти гаметы являются гаметами
родительского типа. В этом случае частота
кроссинговера будет равна 100% –
(2
частота гамет ab).
Задача
1. Пусть
гены А
и В
сцеплены и расстояние между ними
составляет 40 сМ. Сколько и каких гамет
будет образовываться у особей с генотипом
AB
и генотипом Ab
?
ab
aB
Решение.
Для
решения этой задачи необходимо помнить,
что расстояние между генами соответствует
частоте кроссинговера. Частота же
кроссинговера определяется как доля
кроссоверных гамет. Теперь, согласно
этим положениям, мы можем выписать все
типы гамет и указать частоту их
образования.
Для
особи с генотипом AB
:
ab
|
Тип гамет |
Класс гамет |
Частота |
|
Родительский |
AB |
30 |
|
Ab |
30 |
|
|
Кроссоверный |
aB |
20 |
|
Ab |
20 |
Для
особи с генотипом AB
:
ab
|
Тип гамет |
Класс гамет |
Частота |
|
Родительский |
Ab |
30 |
|
aB |
30 |
|
|
Кроссоверный |
AB |
20 |
|
ab |
20 |
О
A B
a
b
твет. Для особи
с генотипом : родительские гаметы
AB:
a
A B
a
b
b (30%) и (30%);
кроссоверные гаметы Ab
(20%) и aB
(20%). Для особи с генотипом : родительские
гаметы Ab
(30%) и aB
(30%); кроссоверные гаметы AB
(20%) и ab
(20%).
Задача
2.
У морских свинок белая окраска шерсти
(w)
рецессивна по отношению к черной окраске
(W),
а волнистые волосы (v)
рецессивны по отношению к прямым (V).
Скрещивается гомозиготная морская
свинка, имеющая белую окраску шерсти и
волнистые волосы, с морской свинкой,
имеющей черные прямые волосы. Полученные
в результате такого скрещивания гибриды
F1
возвратно скрещиваются с двойным
рецессивом и дают следующее потомство:
30 черных морских свинок с прямыми
волосами, 10 – черных с волнистыми
волосами, 12 – белых с прямыми волосами
и 31 – белая с волнистыми волосами (всего
83 особи).
Наследуются ли
гены, контролирующие у морской свинки
окраску шерсти и форму волоса, независимо?
Если гены сцеплены и между ними происходит
кроссинговер, то с какой частотой это
случается?
Решение.
Предположим, что
гены, контролирующие у морской свинки
окраску шерсти и форму волоса, действительно
наследуются независимо друг от друга.
Тогда мы должны получить следующий
результат:
P:
♀
WWVV
♂
wwvv
F1:
WwVv
P:
♀
WwVv
♂
WwVv
Fa:
1/4 черных с прямыми волосами (WwVv);
1/4
черных с волнистыми волосами (Wwvv);
1/4
белых с прямыми волосами (wwVv);
1/4
белых с волнистыми волосами (wwvv).
Проверим
наше предположение с помощью методы
2.
Поскольку в анализирующем скрещивании
было получено всего 83 особи, то в случае
независимого наследования каждый
фенотипический класс должен состоять
из 20,75 особей (1/4
83 = 20,75). Тогда
2факт
= ((30 — 20,75)2/20,75)
+ ((10 — 20,75)2/20,75)
+ ((12 — 20,75)2/20,75)
+ ((31 — 20,75)2/20,75)
= 118,44
df
= n
— 1 = 4 — 1 = 3
20,05
= 7,815
2факт
>> 20,05.
Эти данные указывают
на то, что наше предположение неверно.
Следовательно, гены, контролирующие у
морской свинки окраску шерсти и форму
волоса, сцеплены, но их сцепление не
полное. На основании этого вывода запишем
окончательную схему скрещивания:
P:
♀
W
V
♂ w
v
W V
w v
F1:
W
V
w
v
P:
♀
W
V
♂ w
v
w
v
w
v
Fa:
30 черных с прямыми волосами (некроссоверные
потомки с генотипом WwVv);
31 белая с волнистыми
волосами (некроссоверные потомки с
генотипом Wwvv);
10 белых с волнистыми
волосами (кроссоверные потомки с
генотипом wwVv);
12 белых с прямыми
волосами (кроссоверные потомки с
генотипом wwvv).
Найдем
частоту кроссинговера между генами W
и V:
сумма
кроссоверных особей
rf
=
100%
сумма
всех особей
rf
= ((10 + 12)/(30 + 10 + 12 + 31))
100% = 26,5%
Ответ.
Гены W
и V
сцеплены, но сцепление не полное: rf
= 26,5%.
Задача
3.
При скрещивании душистого горошка,
имеющего яркую окраску цветов и усики
на листьях с растением с бледной окраской
цветков и без усиков на листьях, было
получено первое поколение, состоящее
из растений с яркими цветками и усиками
на листьях. Во втором поколении этого
скрещивания было получено расщепление:
424 растения с яркими цветками и усиками,
99 — бледных с усиками, 102 –ярких без
усиков, 91 – бледных без усиков. Всего:
716 растений. Объясните полученные
результаты.
Решение.
Согласно
условию задачи, при скрещивании душистого
горошка, имеющего яркую окраску цветов
и усики на листьях с растением с бледной
окраской цветков и без усиков на листьях,
в первом поколении было получено
единообразие. Следовательно, исходные
растения были гомозиготными. На основании
этого мы можем ввести обозначения
аллелей, отвечающих за тот или иной
признак: А
– яркая окраска цветков, а
– бледная окраска цветков, В
– наличие усиков и b
– отсутствие усиков.
Если
бы гены наследовались независимо, то
во втором поколении было бы расщепление
9/16 A—B—,
3/16 A—bb,
3/16 aaB—
и 1/16 aabb.
Однако полученные результаты не
соответствуют такому расщеплению, что
подтверждается с помощью метода 2
(при независимом наследовании 2
= 65 при p
0,05). Следовательно, гены сцеплены, но
сцепление не полное.
 Частоту
Частоту
кроссинговера (силу сцепления) можно
определить по частоте особей с генотипом
aabb,
которые возникают при слиянии двух
одинаковых гамет (при этом частоты гамет
перемножаются). Частота образования
особей aabb
= 91:716 = 0,13. Отсюда частота образования
гаметы ab
=
0,13
0,36 или 36%. Поскольку гамета ab
возникает с частотой
36% (т.е. более 25%), то она является
родительской. Точно с такой же вероятностью
образуется гамета АВ.
Следовательно, в сумме доля двух
родительских гамет составляет 72%. Отсюда
частота кроссинговера равна 100% — 72% =
28%. Доля кроссоверных гамет равна 28%.
Ответ.
Гены, контролирующие у душистого горошка
интенсивность окраски цветков и наличие
усиков, сцеплены, но сцепление не полное.
Частота кроссинговера между этими
генами равна 28%.
ЗАДАЧИ ДЛЯ
САМОСТОЯТЕЛЬНОГО РЕШЕНИЯ
-
П
A
bA
ba
Ba
Bусть гены А
и В
сцеплены и частота рекомбинации между
ними составляет 40%. Каковы будут генотипы
особей первого поколения от скрещивания
?
Какие
гаметы и в каком количественном
соотношении будут образовываться у
особей первого поколения? Каким будет
расщепление во втором поколении?
-
Г
A B
a
bены A
и B
сцеплены, кроссинговер между ними
составляет 20%. Определите, сколько
появиться в потомстве дигетерозиготы
при
при
ее самооплодотворении форм с генотипами
aabb
и A—bb?
Что получиться при кроссинговере 10%?
-
Е
a
ba
ba
Ba
B
A
bA
b
a
Ba
Bсли гены А
и В
сцеплены и кроссинговер между ними
составляет 20%, то какова будет пропорция
генотипов и в F2
от скрещивания
? -
У
Drosophila
melanogaster
гены красной, белой и эозиновой окраски
глаз являются разными аллелями одного
гена. Известно, что гены желтого тела
и белых глаз сцеплены друг с другом и
дают 1,5% кроссинговера. Какой процент
кроссинговера будет наблюдаться между
генами желтого тела и эозиновых глаз? -
У
Drosophila
melanogaster
мутация «black»
(черный) отличается черной окраской
тела от нормальных мух, имеющих серую
окраску, а мутация «are»
(дуговидный) в отличие от прямых крыльев
нормальных мух имеет крылья, несколько
вогнутые и загнутые книзу. В ниже
представленном скрещивании указаны
родители (гомозиготы) и потомство,
полученное от скрещивания самок первого
поколения с черными дуговидными самцами
(данные из Бриджеса и Моргана). На
основании этих данных вычислите силу
сцепления между генами, определяющими
окраску тела и форму крыльев.
P:
♀
черное тело
♂
черное тело
дуговидные
крылья дуговидные крылья
F2:
серые,
прямокрылые –
1641,
серые,
дуговидные
– 1251,
черные,
прямокрылые
– 1180,
черные,
дуговидные
– 1532 (всего: 5604).
-
У
Drosophila
melanogaster
признаки желтого тела («yellow»)
и киноварных глаз («vermilion»)
сцеплены с полом и рецессивны по
отношению к серому телу и красным глазам
нормальных особей. Гены желтого тела
и киноварных глаз расположены на
расстоянии 28 сМ. Ген зачаточных крыльев
рецессивен и локализован в одной из
аутосом. Скрещена гомозиготная желтая
красноглазая длинокрылая самка с
гомозиготным серым киноварным
зачаточнокрылым самцом. Каким будет
потомство от скрещивания а) самки из
первого поколения с желтым киноварным
зачаточнокрылым самцом? б) самца из
первого поколения с желтой киноварной
зачаточнокрылой самкой?
-
Джонс
нашел, что у помидора высокий рост
стебля доминирует над карликовым, а
шаровидная форма плода — над грушевидной.
Гены, определяющие эти признаки сцеплены
и частота рекомбинации между ними
соответствует 20%.
Скрещено
гомозиготное высокое растение, имеющее
грушевидные плоды, с гомозиготным
карликовым, имеющим шаровидные плоды.
Каковы будут результаты первого и
второго поколений? Каковы будут результаты
возвратного скрещивания F1
c
карликовым грушевидным?
-
У
R
PR
Pкрыс темная окраска глаз обусловлена
комплиментарным взаимодействием генов
R
и P;
рецессивные аллели каждого из этих
генов вызывают светлую окраску глаз.
Гены эти лежат в одной хромосоме.
r
p
r
p
R
p
R
p
r
P
r
P
Крысы с темными глазами
были скрещены с двойным рецессивом
. При возвратном скрещивании особей
первого поколения с двойными рецессивными
получено следующее потомство (данные
Кастла): темноглазых – 1255, светлоглазых
– 1777.
Когда
же были скрещены крысы с крысами
и F1
возвратно скрестили с двойным рецессивом,
то получилось следующее потомство:
темноглазых – 174, светлоглазых – 1540.
Всего: 1714. Определите частоту рекомбинации
между генами R
и P.
-
Пусть
у кур гены E
(раннее оперение) и B
(рябое оперение) сцеплены с полом и
расстояние между ними 20 сМ. Каково будет
потомство от скрещивания рано оперившейся
черной курицы с петухом, полученным от
скрещивания гомозиготного поздно
оперившегося рябого петуха с рано
оперившейся черной курицей? -
Особь,
гомозиготная по генам A
и B,
скрещена с особью, гомозиготной по
генам a
и b.
Во втором поколении от этого скрещивания
получено: AB
– 334, Ab
– 37, aB
– 38 и ab
– 87 особей. Всего: 496 особей. Чем отличаются
эти результаты от ожидаемых в случае
независимого наследования? Какова
частота рекомбинации между генами А
и В? -
Окраска
зерен у пшеницы обусловлена взаимодействием
двух генов по типу некумулятивной
полимерии. Каково будет расщепление
во втором поколении от скрещивания
красной и белой пшеницы, если расстояние
между генами, определяющими окраску,
составляет 10 сМ? -
C
BC
bD
fd
FСцепленные гены С
и В
дают 20%, а D
и F
— 40% кроссинговера. Гены С
и В
лежат в одной хромосоме, а D
и F
— в другой. Какие и с какой частотой
будут образовываться гаметы у особи с
генотипом . -
Проводиться
анализирующее скрещивание дигетерозиготного
растения AaBb
с полным рецессивом aabb.
Известно, что эти два гена сцеплены, но
сцепление не полное: между ними возможен
кроссинговер с частотой 10%. Кроме того,
известно, что у дигетерозиготной особи
доминантные аллели находятся в
цис-положении. Установите, какую долю
потомства от такого скрещивания будут
составлять особи с генотипом AaBb? -
Скрещивается две
гомозиготные особи EEFF
и eeff.
Полученные в результате такого
скрещивания гибриды F1
возвратно скрещиваются с двойным
рецессивом и дают следующее потомство:
2/6 EF,
1/6 Ef,
1/6 eF
и 2/6 ef.
Объясните эти результаты. -
Скрещивается две
гомозиготные особи AABB
и aabb.
Полученные в результате такого
скрещивания гибриды F1
возвратно скрещиваются с двойным
рецессивом. Какая доля потомства FB
будет иметь генотип aabb,
при условии, что эти два гена а) не
сцеплены; б) полностью сцеплены; в)
располагаются на расстоянии 10 сМ; г)
располагаются на расстоянии 24 сМ? -
Штамм
Neurospora
с генотипом HI
скрещивается со штаммом, имеющим генотип
hi.
Половина потомства от такого скрещивания
имеет генотип HI,
а вторая половина – hi.
Объясните полученный результат. -
У
некоего гаплоидного организма два гена
C
и D
сцеплены и находятся на расстоянии 8
сМ. Проводиться скрещивание типа Cd
cD.
Какую долю от общего количества потомков
от такого скрещивания будут иметь особи
а) CD;
б) cd;
в) Cd;
г) все кроссоверные? -
У
человека дальтонизм и гемофилия
обусловлены сцепленными с полом
рецессивными генами d
и h
(их нормальные аллели – D
и H).
У женщины 6 сыновей: двое из них страдают
дальтонизмом, но имеют нормальную
свертываемость крови; трое страдают
гемофилией при нормальном зрении; один
страдает и гемофилией и дальтонизмом.
Каков генотип матери? Почему она имеет
сыновей трех типов? Могут ли у такой
женщины родиться здоровые сыновья? -
У
человека локус резус-фактора сцеплен
с локусом, определяющим форму эритроцитов,
и находиться от него на расстоянии 3
сМ. Резус-положительность и эллиптоцитоз
(эритроциты эллиптической формы)
определяются доминантными аутосомными
генами. Один из супругов гетерозиготен
по обоим генам. При этом резус-положительность
он унаследовал от одного родителя,
эллиптоцитоз – от другого. Второй
супруг резус-отрицателен и имеет
нормальные эритроциты. Определите
процентное соотношение вероятных
генотипов и фенотипов детей в этой
семье. -
У
человека синдром дефекта ногтей и
коленной чашечки определяется доминантным
аутосомным геном. На расстоянии 10 сМ
от него находиться локус групп крови
системы АВО.
Один из супругов имеет II
группу крови, другой – III.
Тот, у которого II
группа крови, страдает дефектом ногтей
и коленной чашечки. Известно, что его
отец был с I
группой крови и не имел этих аномалий,
а мать – с IV
группой крови и имела оба дефекта.
Супруг, имеющий III
группу крови, нормален в отношении гена
дефекта ногтей и коленной чашечки и
гомозиготен по обеим парам анализируемых
генов.
Определите
вероятность рождения в такой семье
детей, страдающих дефектом ногтей и
коленной чашечки, и возможные у них
группы крови.
-
У
человека рецессивный ген с
обуславливает цветовую слепоту
(протанопию), а рецессивный ген d
– мышечную дистрофию Дюшена. Оба
признака наследуются сцеплено с полом.
По родословной одной многодетной семьи
были получены следующие данные: здоровая
женщина с нормальным зрением, отец
которой страдал мышечной дистрофией,
а мать – нарушением цветового зрения,
вышла замуж за здорового мужчину с
нормальным цветовым зрением. От этого
брака родилось 8 мальчиков и 3 девочки.
Из них три девочки и один мальчик были
здоровы и имели нормальное зрение. Из
остальных семи мальчиков 3 страдали
мышечной дистрофией, 3 – цветовой
слепотой и 1 – обоими заболеваниями.
По этим данным была дана приблизительная
(ввиду малочисленности материала)
оценка расстояния между генами c
и d.
Укажите это расстояние. -
Бешеный
огурец является двудомным растением,
причем пол у него определяется не
морфологически различимыми половыми
хромосомами, а благодаря двум генам.
Контроль формирования пола со стороны
этих генов происходит следующим образом:
один из генов, ген М,
детерминирует мужскую фертильность,
а его рецессивная аллель m
– мужскую стерильность; второй ген,
ген F,
определяет женскую стерильность, а его
рецессивная аллель f
– женскую фертильность. Любая популяция
бешенного огурца состоит из мужских
(примерно 50%) и женских (так же примерно
50%) особей. Кроме того, очень редко в
таких популяциях встречаются
гермафродитные особи, которые имеют
как мужские, так и женские половые
органы.
Каковы полные
генотипы мужских и женских особей у
бешенного огурца? Благодаря чему в
популяциях такого растения поддерживается
примерно равное соотношение мужских и
женских особей? Какова причина появления
у бешенного огурца гермафродитных
особей и почему они появляются с очень
низкой частотой?
-
У
льна известны линии, которые по-разному
чувствительны к некоторым расам льняной
ржавчины. Например, линия льна 77ОВ
резистентна к ржавчине расы 24, но
восприимчива к расе 22. Линия же Бомбей,
наоборот, к расе 24 восприимчива, но зато
устойчива по отношению к расе льняной
ржавчины 22. Если скрестить эти две
линии, то гибриды F1
будут устойчивы к обеим расам
фитопатогенного гриба. При дальнейшем
же размножении гибридов F1
все потомство F2
распределяется следующим образом:
растений, резистентных к обеим расам
льняной ржавчины – 184, резистентных к
расе 22, но чувствительных к расе 24 –
58, резистентных к расе 24, но чувствительных
к расе 22 – 63, чувствительных к обеим
расам гриба – 15. Предложите гипотезу,
которая могла бы объяснить эти данные.
Подтвердите или опровергните правильность
выдвинутой гипотезы с помощью метода
2.
Укажите генотипы исходных линий льна
77ОВ и Бомбей, генотип гибридов F1
и генотипы всех потомков F2. -
У
наземной улитки Cepea
nemoralis
цвет раковины определяется тремя
аллелями одного гена: Y
– коричневый, y1
– розовый, y
– желтый. Аллель Y
доминирует над аллелями y1
и y,
а аллель y1
– над аллелью y.
С локусом окраски тесно сцеплен ген
распределения окраски (полосатости).
Этот ген так же представлен тремя
аллелям: аллель B
контролирует равномерное распределение
коричневой, розовой и желтой окраски,
аллель b1
детерминирует появление широкой темной
полосы на общем фоне, а аллель b
– несколько узких полос. Степень
доминирования среди этих аллелей
следующая: B
> b1
> b.
Соседние файлы в предмете [НЕСОРТИРОВАННОЕ]
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
- #
Как уже отмечалось, Г. Мендель, установивший важнейшие закономерности наследования, выдвинул предположение о существовании особых наследственных факторов, контролирующих признаки живых организмов. Однако материальная природа этих факторов, впоследствии названных генами, долгое время оставалась невыясненной. Лишь в начале ХХ в., после переоткрытия законов Г. Менделя, цитологи обратили внимание на связь гипотетических наследственных факторов с поведением хромосом при мейозе и оплодотворении. В 1902—1903 гг. немецкий эмбриолог Т. Бовери и американский цитолог У. Саттон (фамилии приведены не для запоминания) независимо друг от друга пришли к выводу, что именно хромосомы и являются носителями менделевских факторов. Однако ученые не смогли предоставить четких доказательств своей правоты, поэтому их предположение в течение нескольких лет так и оставалось «хромосомной гипотезой».
Хромосомная теория наследственности. Сцепленное наследование. Убедительные доказательства того, что гены располагаются в хромосомах, были получены в 1910 г. американским генетиком, будущим лауреатом Нобелевской премии (1933 г.) Т. Морганом. Многочисленные эксперименты Моргана и его сотрудников привели к ряду важнейших открытий, которые легли в основу хромосомной теории наследственности. Одно из положений этой теории можно сформулировать следующим образом: гены расположены в хромосомах в линейном порядке и занимают определенные участки — локусы, причем аллельные гены находятся в одинаковых локусах гомологичных хромосом.
Закон независимого наследования (третий закон Менделя) справедлив только в том случае, если неаллельные гены находятся в разных парах хромосом. Однако количество генов у живых организмов значительно больше числа хромосом. Например, у человека около 25 тыс. генов, а количество хромосом — 23 пары (2n = 46), у плодовой мушки дрозофилы приблизительно 14 тыс. генов и всего 4 пары хромосом (2n = 8). Следовательно, каждая хромосома содержит множество генов. Будут ли гены, локализованные в одной хромосоме, наследоваться независимо? Очевидно, что нет.
Гены, расположенные в одной хромосоме, образуют группу сцепления и наследуются вместе. Такое совместное наследование генов Т. Морган назвал сцепленным наследованием (в отличие от независимого). Гомологичные хромосомы каждой пары содержат гены, контролирующие одни и те же признаки, поэтому количество групп сцепления равно числу пар хромосом. Например, у человека 23 группы сцепления, а у дрозофилы — 4.
Вам известно, что при независимом наследовании дигетерозиготная особь, например , образует четыре типа гамет в равном соотношении, т. е. по 25 %:
,
,
и
. Это обусловлено тем, что неаллельные гены находятся в разных парах хромосом. Если же они расположены в гомологичных хромосомах, следовало бы ожидать, что дигетерозигота
будет производить лишь два типа гамет: 50 %
и 50 %
. Обратите внимание на то, что сцепленные гены записываются в одну хромосому.
Однако Т. Морган обнаружил, что в большинстве случаев дигетерозиготные особи образуют не два типа гамет, а четыре. Кроме ожидаемых и
, формируются также гаметы с новыми комбинациями генов:
и
, но в меньшем процентном соотношении, чем при независимом наследовании. Рассмотрим один из опытов, в котором Т. Морган изучал наследование сцепленных генов у дрозофилы.
 *Дрозофила обыкновенная, или плодовая, — вид насекомых отряда Двукрылые (рис. 34.1). Особи, как правило, имеют желто-коричневое брюшко с темными поперечными кольцами и глаза красного цвета. В природе эти мушки питаются опавшими фруктами, соком растений, гниющими растительными остатками. В XX в. дрозофила стала одним из самых распространенных объектов исследований в области генетики и биологии индивидуального развития. Удобство ее использования обусловлено рядом причин. Так, дрозофилы имеют множество отчетливо различимых альтернативных признаков, небольшие размеры (обычно около 2—2,5 мм), легко разводятся в лабораторных условиях и обладают высокой плодовитостью — от каждого скрещивания можно получить до нескольких сотен потомков. Важно также и то, что время развития особи от откладки яйца до выхода половозрелой мушки из куколки при 25 °C занимает всего 10 дней.*
*Дрозофила обыкновенная, или плодовая, — вид насекомых отряда Двукрылые (рис. 34.1). Особи, как правило, имеют желто-коричневое брюшко с темными поперечными кольцами и глаза красного цвета. В природе эти мушки питаются опавшими фруктами, соком растений, гниющими растительными остатками. В XX в. дрозофила стала одним из самых распространенных объектов исследований в области генетики и биологии индивидуального развития. Удобство ее использования обусловлено рядом причин. Так, дрозофилы имеют множество отчетливо различимых альтернативных признаков, небольшие размеры (обычно около 2—2,5 мм), легко разводятся в лабораторных условиях и обладают высокой плодовитостью — от каждого скрещивания можно получить до нескольких сотен потомков. Важно также и то, что время развития особи от откладки яйца до выхода половозрелой мушки из куколки при 25 °C занимает всего 10 дней.*
Чистую линию дрозофил, имеющих серое тело и нормальные (длинные) крылья, скрестили с чистой линией, особи которой имели черное тело и зачаточные крылья (рис. 34.2). Полученные гибриды первого поколения в соответствии с первым законом Менделя были единообразными — серыми с нормально развитыми крыльями. Следовательно, у дрозофилы серое тело (А) полностью доминирует над черным (а), а нормальные крылья (В) — над зачаточными (b). Все гибриды первого поколения — дигетерозиготы.

Затем было проведено анализирующее скрещивание (рис. 34.3). Дигетерозиготную самку из гибридного поколения скрестили с рецессивным дигомозиготным самцом (черное тело и зачаточные крылья). В потомстве было получено по 41,5 % особей с серым телом, нормальными крыльями и черным телом, зачаточными крыльями, а также по 8,5 % мух с серым телом, зачаточными крыльями и черным телом, нормальными крыльями.

Если бы гены, определяющие цвет тела и развитие крыльев, находились в разных парах хромосом, соотношение фенотипических классов было бы равным — по 25 %. Но этого не наблюдалось, значит, гены находятся в гомологичных хромосомах и наследуются сцепленно.
Несмотря на сцепление генов, самка производила не два, а четыре типа гамет. Однако гамет с исходными сочетаниями сцепленных генов формировалось намного больше (
и
вместе составили 83 %), чем с новыми их комбинациями (сумма
и
равна 17 %).
Было выяснено, что причиной появления хромосом с новыми комбинациями родительских генов является кроссинговер. Вы знаете, что этот процесс происходит в профазе I мейоза и представляет собой обмен соответствующими участками между гомологичными хромосомами. Таким образом, кроссинговер препятствует полному (абсолютному) сцеплению генов. Гаметы, которые образуются в результате кроссинговера, и особи, которые развиваются при участии таких гамет, называются кроссоверными или рекомбинантными. В рассмотренном эксперименте гаметы и
являлись кроссоверными, а гаметы
и
— некроссоверными (см. рис. 34.3).
*Т. Морган подвергал анализирующему скрещиванию не только дигетерозиготных самок дрозофилы, но и самцов, т. е. проводил реципрóкные скрещивания. Так называют пару скрещиваний, в одном из которых организмы с исследуемыми признаками (или признаком) используются в качестве материнских, а в другом — в качестве отцовских. Оказалось, что реципрокные анализирующие скрещивания дигетерозиготных особей дрозофилы дают разные результаты.
В потомстве дигетерозиготного самца наблюдалось лишь два фенотипических класса (рис. 34.4). Половину потомков составляли особи с серым телом и нормальными крыльями, вторую половину — особи с черным телом и зачаточными крыльями. Это свидетельствовало об отсутствии кроссинговера между сцепленными генами или, иначе говоря, о полном (абсолютном) сцеплении генов. Выяснилось, что у самцов дрозофилы в ходе мейоза действительно не происходит кроссинговер между гомологичными хромосомами. Впоследствии подобное явление было обнаружено у самцов некоторых других видов двукрылых, а также у самок тутового шелкопряда. Однако полное сцепление генов наблюдается в природе очень редко, как исключение из общего правила.*

Кроссинговер между сцепленными генами происходит с определенной вероятностью (частотой). Для расчета частоты кроссинговера (rf, от англ. recombination frequency — частота рекомбинации) можно пользоваться следующей формулой:
.
Таким образом, между генами А и В, контролирующими цвет тела и длину крыльев дрозофилы, кроссинговер происходит с частотой: rfAB = 17 %.
Дальнейшие исследования, проведенные Т. Морганом и сотрудниками его лаборатории, показали, что частота кроссинговера пропорциональна расстоянию между генами, расположенными в одной хромосоме. Чем больше расстояние между сцепленными генами, тем чаще между ними происходит кроссинговер. И наоборот, чем ближе друг к другу расположены гены, тем меньше частота кроссинговера между ними. Чем объясняется эта закономерность?
В профазе I мейоза при конъюгации гомологичных хромосом взаимный обмен участками между хроматидами осуществляется произвольно, в любой их точке. Рассмотрим рисунок 34.5.

Гены А и В (или а и b) находятся сравнительно близко друг к другу. Вероятность того, что обмен произойдет именно на участке, разделяющем эти гены, невелика. Гены А и D (или а и d) располагаются на значительном расстоянии друг от друга. Поэтому вероятность того, что хроматиды совершат обмен в какой-либо точке между этими генами, намного выше. Значит, чем больше расстояние между генами, тем чаще они разделяются при кроссинговере.
Таким образом, частота кроссинговера позволяет судить о расстоянии между генами. В честь Т. Моргана единица измерения расстояния между генами получила название морганида или, что то же самое, сантиморган (сМ).
Морганида (сантиморган, сМ) — это генетическое расстояние, на котором кроссинговер происходит с вероятностью 1 %.
Кроссинговер играет очень важную биологическую роль. В результате этого процесса у потомков возникают новые комбинации родительских генов, что повышает генетическое разнообразие организмов и расширяет возможности их адаптации к различным условиям окружающей среды.
Генетические карты. Как показали эксперименты Т. Моргана, результаты анализирующего скрещивания дают возможность определять частоту кроссинговера между сцепленными генами и судить о расстоянии между ними *(за исключением тех редких случаев, когда кроссинговер отсутствует)*. Использование данных, полученных от множества анализирующих скрещиваний с различными комбинациями сцепленных генов, позволяет ученым составлять так называемые генетические карты (рис. 34.6). Генетическая карта хромосомы — это схема взаимного расположения генов, находящихся в одной группе сцепления, построенная с учетом расстояний между ними. Такие карты уже составлены для человека и многих других видов живых организмов, например для мыши, дрозофилы, гороха, пшеницы, кукурузы, томата, дрожжей и т. д.
.

Генетические карты находят широкое применение в научных исследованиях и практической деятельности человека. Знание генетических карт позволяет предсказывать характер наследования тех или иных признаков. Использование этих данных в селекции и генетической инженерии способствует планированию работы по получению организмов с определенными сочетаниями признаков. Сравнение генетических карт разных видов организмов дает возможность устанавливать степень их родства, судить о возможных путях эволюционных преобразований наследственного материала.

Исследования Т. Моргана и его сотрудников легли в основу хромосомной теории наследственности, основными положениями которой являются следующие.
1. Гены в хромосомах расположены линейно, в определенной последовательности. Аллельные гены находятся в одинаковых локусах гомологичных хромосом.
2. Гены, расположенные в одной хромосоме, образуют группу сцепления и наследуются вместе. Количество групп сцепления равно числу пар хромосом.
3. Сцепление генов может нарушаться в результате кроссинговера, происходящего при конъюгации гомологичных хромосом в профазе I мейоза.
4. Частота кроссинговера пропорциональна расстоянию между генами: чем больше расстояние, тем выше частота кроссинговера и наоборот.
5. За единицу расстояния между сцепленными генами принята 1 морганида (1 сантиморган, сМ). Это расстояние, на котором кроссинговер происходит с вероятностью 1 %.
Схемы взаимного расположения генов, находящихся в одной группе сцепления, построенные с учетом расстояний между ними, называют генетическими картами хромосом. Такие карты используются в научных исследованиях и различных сферах практической деятельности человека.
 |
1. Всегда ли соблюдается III закон Г. Менделя? В каких случаях гены наследуются независимо, а в каких — сцепленно? 2. Почему частота кроссинговера между сцепленными генами зависит от расстояния между ними? 3. Что представляют собой генетические карты хромосом? Для чего они используются? 4. Сформулируйте основные положения хромосомной теории наследственности. 5. Какие типы гамет и в каком процентном соотношении будут формировать дигетерозиготные особи 6*. Серый цвет тела у дрозофилы доминирует над желтым, красные глаза — над гранатовыми. Гены, отвечающие за эти признаки, локализованы в первой паре хромосом и находятся на расстоянии 44 морганиды. Скрестили чистые линии серотелых мух с гранатовыми глазами и желтотелых с красными глазами. Из полученных гибридов выбрали самку и подвергли ее анализирующему скрещиванию. Каким будет процентное соотношение фенотипических классов в потомстве? 7*. У мака высокий стебель (А) доминирует над низким, крупные листья (В) — над мелкими, красные цветки (С) — над белыми. С тремя различными дигетерозиготными растениями были проведены анализирующие скрещивания, результаты которых представлены в таблице. Определите расстояния между генами А, В и С. Постройте генетическую карту хромосомы мака, в которой локализованы эти гены.
|